| Edukir hat folgendes geschrieben: |
|
Heute will ich darstellen, wie man sich aus Sicht der Armed Ape Theory der Frage nach der Größe des menschlichen Gehirns annähern müßte. Obwohl die Gehirngröße im Gegensatz zu den Anpassungen an das gezielte Werfen schon immer im Mittelpunkt der wissenschaftlichen Diskussion gestanden hat, wirken die bislang präsentierten Erklärungsmuster eher hilflos. |
| Edukir hat folgendes geschrieben: |
|
Manche Wissenschaftler verbreiten den Standpunkt, dass es keinen Zusammenhang zwischen Gehirngröße und Leistungsfähigkeit des Gehirns beim Menschen gäbe. Nicht die Größe, sondern die "Verdrahtung" soll entscheidend für die Leistungsfähigkeit des Gehirns sein. Ergibt dieser Standpunkt einen Sinn? Ich denke das tut er nicht. Ein größeres Gehirn geht mit enormen fitnessrelevanten Kosten einher. Es verbraucht mehr Energie, es bereitet Schwierigkeiten bei der Geburt. Es zwingt Frauen dazu sehr hilfsbedürftige Babys zur Welt zu bringen und mehr in die Aufzucht dieser Kinder zu investieren. Es verlängert die Generationendauer und geht damit zu Lasten der Reproduktionsrate. Große Gehirne mußten schon einiges leisten, um all diese Kosten rechtfertigen zu können - sonst hätten sie sich im Rahmen der Evolution schlicht nicht entwickelt. |
| Edukir hat folgendes geschrieben: |
|
Nun könnte man einwenden, dass es dann auch einen nachweisbaren Zusammenhang zwischen Gehirngröße und Gehirnleistungen beim Menschen geben müßte, ein solcher sei aber weit und breit nicht in Sicht. Dem ersten Argument stimme ich zu - dem zweiten nicht. |
| Edukir hat folgendes geschrieben: |
|
Welche Hinweise stehen uns konkret zur Verfügung, wenn wir uns auf die Suche nach einem Leistungsmerkmal machen wollen, das beim Menschen mit der Gehirngröße korreliert sein könnte? Wir verfügen über eine ganze Reihe von Indizien, die uns Hinweise darauf geben, wo wir mit der Suche anfangen sollten: 1) Schädelfunde geben uns Hinweise, wann das Gehirn im Laufe der menschlichjen Evolution vergrößert wurde. So richtig los ging dieser Prozess vor knapp 2 Millionen Jahren. Mit der Entwicklung moderner Körperproportionen beim Übergang zum Homo erectus vor ca. 1.8 Millionen Jahren kam auch das Gehirnwachstum richtig in Fahrt. Wir suchen also nach einer aus Sicht des Gehirns sehr anspruchsvollen Tätigkeit, die bereits vor 1.8 Millionen Jahren zum Verhaltensrepertoire unserer Vorfahren zählte und am Besten auch gleich einen Beitrag zur Klärung der Frage leisten könnte, warum Unsere Vorfahren den menschlichen Körperbau entwickelt haben. |
| Edukir hat folgendes geschrieben: |
|
2) Männer haben größere Gehirne als Frauen. Ein deutlicher Sexualdimorphismus bei der Gehirngröße ist von vorn herein zu beobachten - bei den 1.8 Millionen Jahre alten Dmanissi-Homininen steht zwei vermutlich weiblichen Gehirnkapazitäten von 600 und 650 ccm eine vermutlich männliche von 770 ccm gegenüber. Bei den Neandertalern hatten Männer eine um etwa 200 ccm höhere Gehirnkapazität als Frauen. es liegt daher nahe nach einem Leistungsmerkmal zu suchen, bei dem Männer im Vergleich zu Frauen deutlich überlegen sind. |
| Edukir hat folgendes geschrieben: |
|
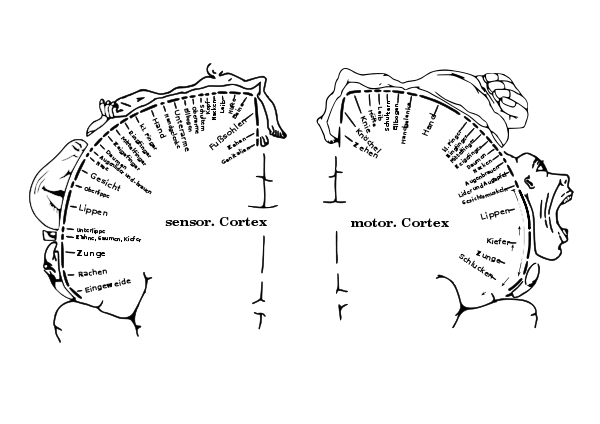
3) Gehirne sind bei Männern nicht nur im Durchschnitt größer, sie sind auch funktional stärker lateralisiert. Es liegt daher nahe, nach einer Tätigkeit zu suchen, bei der eine deutliche Arbeitsteilung der beiden Gehirnhälften zu einer höheren Leistungsfähigkeit führt. Das war vermutlich nicht die Sprachfähigkeit, weil Frauen, bei denen die Sprachfunktionen weniger lateralisiert sind, Männern in sprachlicher Hinsicht keineswegs unterlegen sind. Vielversprechender ist es wohl nach einer ausgesprochen asymmetrischer Tätigkeit zu suchen, bei der der dominanten Hand eine Schlüsselrolle zukommt. Für eine zentrale Bedeutung der Hand für die stärkere Lateralisierung des männlichen Gehirns spricht die Händigkeit und die starke Repräsentanz der Hand im primären motorischen und sensorischen Kortex. |
| Edukir hat folgendes geschrieben: |
|
4) Sowohl bei der Entwicklung moderner Menschen in Afrika als auch der Neandertaler in Europa sind die Gehirne weiter angewachsen und wiesen am Ende in etwa die gleiche Größe auf. Wir suchen also nach einem Verhaltensmerkmal, das unter sehr unterschiedlichen klimatischen Bedingungen seine Wirkung entfaltete. Damit fallen z.B. die Jagd odar das Sozialverhalten durchs Raster. Die Jagd war für die Neandertaler wesentlich wichtiger und hätte in Europa zur Entwicklung der größeren und leistungsfähigeren Gehirne führen müssen. Die Komplexität des Sozialverhaltens hängt wiederum mit der Gruppengröße zusammen, die bei den Neandertalern aus ökologischen Gründen deutlich limitiert war. Hätte das Sozialverhalten eine Schlüsselrolle bei der Gehirnexpansion gespielt, dann wäre die Gehirngröße bei den Neandertalern deutlich zurückgeblieben. |
| Edukir hat folgendes geschrieben: |
|
Gibt es eine aus Sicht des Gehirns sehr anspruchsvolle Tätigkeit, die in dieses Suchraster passt? Ja, es ist das gezielte Werfen. |

| Edukir hat folgendes geschrieben: |
|
Was ist also aus Sicht der Armed Ape Theory der menschlichen Evolution zu tun, wenn man klären will, warum das menschliche Gehirn so groß ist? - Fürs Erste muß man versuchen festzustellen, ob es eine Korrelation zwischen Gehirngröße und Leistungsfähigkeit beim gezielten Werfen gibt. Dazu nimmt man am besten eine statistisch relevante Zahl von Spitzensportlern, die sich als besonders Leistungsfähig im gezielten Werfen erwiesen haben (Baseballspieler, Handballspieler etc.) und vergleicht deren durchschnittliche Gehirngröße mit der durchschnittlichen Gehirngröße einer Kontrollgruppe. Ich wette um eine Kiste Bier, dass der Durchschnittswert bei den Werfern um mindestens 100 ccm höher liegen wird. Sollte ich mit dieser Vorhersage recht behalten, dann wäre es an der Zeit für einen Paradigmenwechsel in den beteiligten Wissenschaftsdisziplinen. Der Mensch ist dann nicht als "unspezialisiertes Wesen" aufzufassen, sondern als ein in höchstem Maße spezialisierter Primat. Seine Spezialisierung wäre dann identifiziert und könnte eingehend untersucht werden. Entscheidend ist die Werfer-Hypothese nicht einfach nur als interessante Idee zu konsumieren, sondern sie tatsächlich auf die Probe zu stellen. |
| Lamarck hat folgendes geschrieben: |
| .....
Noch einmal: Es gibt keine Anpassungen an Tätigkeiten oder Verrichtungen. Anpassungen betreffen Umweltbedingungen. Warum sollen 'Anpassungen' an ein Werfen 'wertiger' sein als beispielsweise Anpassungen an 'Nasenbohren' oder 'Autofahren'?! Du kannst Dir aber mal überlegen, was Du so den ganzen Tag machst mit Deinen Armen/Händen ... ...... |
| Lamarck hat folgendes geschrieben: |
| Es gibt keinen Zusammenhang. Vgl. etwa:
Schiff, M. & Lewontin, R.C. (1986): Education and Class. Oxford: Oxford University Press. Ich nehme an, wir brauchen uns hier mit so etwas wie "The Bell Curve" nicht auseinandersetzen ... |
| Lamarck hat folgendes geschrieben: | ||
Hi Edukir!
Es zählen: Anzahl der Neuronen, Komplexität der Verschaltung, Organisation. Beim Menschen ergibt sich hierdurch eine geschätzte Speicherleistung von 2 * 10^15 Byte = 2 PB |
| Zitat: | ||
Es gibt keinen Zusammenhang. Vgl. etwa: Schiff, M. & Lewontin, R.C. (1986): Education and Class. Oxford: Oxford University Press. |
| Zitat: | ||
Die anspruchsvollste Tätigkeit eines Gehirns ist Denken, findest Du nicht? Und die wichtigste Fähigkeit des Denkens ist das Erkennen von Irrtümern, oder? |
| Zitat: | ||
Korreliert mit Körpergröße/Körpergewicht. Übrigens spielt hier u. a. auch das Alter eine wesentliche Rolle. |
| Zitat: | ||
Wieso soll die Jagt für den Neandertaler wichtiger gewesen sein als für den Kollegen? Welche ökologischen Gründe sollen hier limitierend gewesen sein? Wie sieht die Sache im Vergleich mit Homo erectus aus? |
| Zitat: | ||
Kimme, Korn und Schuss: |
| Zitat: | ||
Wenn es um die absolute Gehirngröße geht, nehme ich Gewichtheber im Superschwergewicht oder Stabhochspringer. Wenn es auf den relativen Anteil zum Körpergewicht ankommen soll: Bodenturnen (Frauen). Merke: Nicht unbedingt ist derjenige mit dem größten Kopf auch der bessere Denker ... . |
| Norm hat folgendes geschrieben: |
| Aus dem Biounterricht, ist schon ein paar Jährchen her, aber so ungefähr wurde das gelehrt:
- Nicht die Masse des Gehirns ist wichtig, sondern die Größe der Oberfläche (faltige Struktur). - Nur ein Teil der Gehirnzellen wird auch wirklich aktiv genutzt, daneben gibt es auch inaktive Zellen. |
| Ilmor hat folgendes geschrieben: |
| @Edukir
Wirklich eine interessante These. Ich hatte auch schon mal einen ähnlichen Gedanken, allerdings bezogen auf den Speer. Eine Frage, wenn wir tatsächlich evolutionär aufs Werfen getrimmt sein sollten, müssten wir dann nicht den instiktiven Wunsch zum Werfen verspüren, und würde es dann nicht eine viel größere Rolle in unserem Alltagsleben spielen (mehr als das bisschen Sport). |
| Edukir hat folgendes geschrieben: |
| Die Kultur ist eine sehr mächtige Trumpfkarte, wenn es darum geht ein Szenario der Menschwerdung zu entwickeln. |
| Edukir hat folgendes geschrieben: | ||
Diese Hypothese erklärt nicht das große Gehirn - man hätte die zusätzlichen Nahrungsmittel ja auch in zusätzliche Nachkommen stecken können. Es bleibt also dabei, dass die Gehirnleistungen dessen Kosten rechtfertigen müssen. |
| Edukir hat folgendes geschrieben: |
| Auf jeden Fall schreien derartige Leistungen geradezu nach einer Erklärung. Und die müßte im Falle des Werfens ja wohl im Rahmen einer Theorie der Menschwerdung erfolgen. Das heißt natürlich nicht automatisch, dass meine Theorie komplett richtig ist. Aber es besteht in jedem Fall Handlungsbedarf. Ein Zeitfenster von 1 ms verdient meines Erachtens mehr Aufmerksamkeit als ein 400 000 Jahre alter Wurfspeer. |
| smallie hat folgendes geschrieben: | ||
Kultur könnte tatsächlich der Knackpunkt sein. Und zwar "kummulative" Kultur. Es gibt im Tierreich unzählige Beispiele für Kultur. Ich erwähne nur mal Nüsse knacken oder mit Stöcken nach Termiten angeln bei Schimpansen. Weitere Beispiele ließen sich für viele andere Tierarten geben. Aber das sind alles Errungenschaften, die ein Individuum auch erneut alleine "erfinden" könnte, sollte eine Errungenschaft mal verloren gehen. "Kummulative" Kultur gibt es im Wesentlichen nur beim Menschen. Siehe den Spruch von Newton: "If I have seen a little further it is by standing on the shoulders of Giants." Es liegt nahe, daß sich "kummulatives Lernen" und ein größeres/besseres Gehirn gegenseitig bedingen. Erstens, um sich alles merken zu können. Zweitens um Absichten anderer Individuen besser einschätzen zu können, und so über bloßes Lernen durch Nachahmung hinauszugehen. Drittens, später, um Fähigkeiten mittels Sprache besser weiter geben zu können. |
| Zitat: | ||
Allerdings. Hab mal ein bisschen 'rumgesucht und dies hier, ebenfalls von Hore et. al., gefunden: http://jn.physiology.org/cgi/reprint/86/6/2690.pdf Da ist von 7 ms die Rede. Aber auch das wäre immer noch verblüffend. Ich hätte das zeitliche "Aktionsfenster" eher auf 30 - 50 ms geschätzt. Man kann doch nicht schneller handeln, als man wahrnehmen kann? Oder doch? |
| fwo hat folgendes geschrieben: | ||
Weil Werfen dir sowohl Nahrung besorgen als auch Konkurrenten wie Fressfeinde vom Leibe halten kann. Das erfolgreiche Werfen hat damit einen direkteren Einfluss auf deine Reproduktionsfähigkeit als das Nasebohren. Ich weiß jetzt nicht, gegen welche Götter ich gerade argumentiere, aber ich halte das Dogma der Anpassung an Umweltbedingungen in dieser Ausschließlichkeit für Kappes. So nach dem Motto: Und nachdem ein Haufen Erde auf ihn gefallen war und dem Maulwurf dann seine Schaufeln gewachsen waren, begann er zu überlegen, was er damit anfangen sollte.... |
| fwo hat folgendes geschrieben: |
|
Ich nehme an, wir sind uns einig darüber, dass jede Spezialisierung letztendlich in einer wechselseitigen Anpassung = Coevolution von Verhalten und Anatomie stattfinden muss, sonst kämen wir nicht sehr weit. Die Frage ist, was anfängt, und da scheint es mir sinnvoller vom Verhalten auszugehen, weil ein verändertes Verhalten sich am Anfang rein statistisch äußern kann, aber schon dabei automatisch zu einem veränderten Selektionsdruck in der Anatomie führt. |
| fwo hat folgendes geschrieben: |
|
Auch veränderte Umweltbedingungen selektieren Anpassungen an sie nur durch das bereits angepasste Verhalten. Und ein geändertes Verhalten führt auch regelmäßig in / zu andere/n Umweltbedingungen. |
| fwo hat folgendes geschrieben: |
|
Als Beispiel mag der Übergang vom Wasser zum Land dienen: Wie stelle ich mir den vor? Sind da ganz viele Tümpel ausgetrocknet und in einem haben die es geschafft? Mein persönlicher Vorschlag ist, dass da welche vom Schwimmen zum Krauchen übergegangen sind und sich dann über weitere Verhaltensänderungen langsam über die zwangsläufig immer existierenden Uferzonen langsam an Land begeben haben. Ein derartiges Modell gibt der Entwicklung auch mehr Gelegenheit. |

| fwo hat folgendes geschrieben: |
|
Und eigentlich sollte es dir entgegenkommen - gibt es dem Chevalier de Lamarck doch wenigstens über den Umweg der Keimbahn Recht. |
| Messi hat folgendes geschrieben: |
| leer -alae |
| Messi hat folgendes geschrieben: |
|
leer -alae |
| Messi hat folgendes geschrieben: |
|
leer -alae |
| Messi hat folgendes geschrieben: |
|
leer -alae |
| Messi hat folgendes geschrieben: |
|
leer -alae |
| Messi hat folgendes geschrieben: |
|
leer -alae |
| Messi hat folgendes geschrieben: |
|
leer -alae |
| Ilmor hat folgendes geschrieben: |
| @Edukir
Noch eine Frage. Würden beispielsweise die proportional längeren Arme des Schimpansens oder seine uns weit überlegene Oberkörpermuskelkraft nicht einen gewaltigen Vorteil für Werfer darstellen? Warum hat sich der Mensch aber dann in die andere Richtung entwickelt bzw. warum gab es hier keine Rückentwicklung? |
| Zitat: |
| [quote="Lamarck" postid=1576167]
Die Somatotopik bietet doch einigen Aufschluss. Konstruktiv nahezu einzigartig ist der Präzisionsgriff des Menschen, den schon mindestens der Neandertaler auszuführen in der Lage war (Daneben konnte bereits Bambiraptor mit zwei Fingern greifen). Natürlich spielt der Präzisionsgriff beim (einarmigen) Werfen eine Rolle - ist hierzu aber konstruktive Voraussetzung (d. h. erst Präzionsgriff, dann dadurch Potential zum präzisen Werfen). Mithin ist die Fähigkeit zum Werfen konstruktiv ein Abfallprodukt des universellen Präzisionsgriffs - ungeachtet der Tatsache, dass die Fähigkeit zum Werfen Selektionsvorteile bieten kann. |
| Lamarck hat folgendes geschrieben: |
| Es wird also konstatiert, dass bei 'Anpassung' die 'Keimbahn' in einer gewissen Relation mit 'Umwelt' steht. Über die Randbedingung 'Umwelt' ergibt sich somit ein Erklärungswert. Ohne diese "Ausschließlichkeit" ist der Begriff der Anpassung synonym zu einem "Ist so geworden, weil es so geworden ist". |
| kereng hat folgendes geschrieben: | ||
Steht "Th." für "Tidenhub"? |

| Edukir hat folgendes geschrieben: | ||
Wieviel Unsinn willst du hier eigentlich noch verbreiten? Der Präzisionsgriff soll nun die konstruktive Voraussetzung fürs Werfen sein - ungeachtet der Tatsache, daß der Mensch bei weitem nicht der einzige Primat ist, bei dem das Werfen beobachtet werden kann. Nebenbei bemerkt kommt der Präzisionsgriff bei der Verwendung von Wurfspeeren überhaupt nicht zum Einsatz. |


| Lamarck hat folgendes geschrieben: | ||||
Hi Edukir!
Du siehst, ich habe oben etwas hervorgehoben. Primaten sind nicht so gut in der olympischen Disziplin Speerwurf. Zum einen liegt es daran, dass in aller Regel ein Anlauf von einigen Schritten nötig ist, um hier halbwegs einen Blumentopf gewinnen zu können. Dazu ist aber anatomisch schon ein recht ausgefeilter aufrechter Gang mit der Möglichkeit zum komplexen Bewegungsablauf nötig (Der Speer kann zwar auch im Sitzen geworfen werden, das Ergebnis wird allerdings nicht berauschend sein). Gerade der Speerwurf erfordert komplexe Koordination:
Und selbstverständlich kommt der Präzisionsgriff bei der Verwendung von Wurfspeeren zum Einsatz:
Es existieren hier an brauchbaren Grifftechniken:
Ist es nicht verwunderlich, dass Du auch hier eine gewisse Auswahl an Möglichkeiten hast?! Speerwurf mit Fäustlingen scheint allerdings hier nicht sonderlich ratsam ... . Mithin solltest Du nun einigermaßen in der Lage sein, das dargebotene komplexe Zusammenspiel anatomischer Besonderheiten (aufrechter Gang, Präzisionsgriff, Motorik ...) des Menschen evolutiv nachzuvollziehen, nicht? Cheers, Lamarck |
| Edukir hat folgendes geschrieben: |
|
Schön, daß du nun mit Hinweisen auf Werfer - Anpassungen um dich wirfst, [...] |
| Edukir hat folgendes geschrieben: |
|
[...] aber den Präzisionsgriff kann ich dennoch nicht erkennen. Selbst beim Daumen-Mittelfingergriff liegt der Speer nicht zwischen der Fingerkuppe des Zeigefingers und derjenigen des Daumen. Die anderen Finger sind erst recht nicht in Richtung Daumen orientiert. In allen drei Fällen wird der Speer von den Fingern nicht in erster Linie gegen die empfindliche Kuppe des Daumens gedrückt, sondern gegen dessen Basis. Diese drei Griffe stehen dem Kraftgriff näher als dem Feingriff. |
| Edukir hat folgendes geschrieben: |
|
Deine Hervorhebung oben tut auch nichts zur Sache. Das ist nichts anderes, als die alte Behauptung, daß die Evolution prinzipiell nicht in der Lage sei komplexe Strukturen hervorzubringen. |
| Edukir hat folgendes geschrieben: |
|
Richte dich mit deiner Kritik an Evolutionstheoretiker. |

| Edukir hat folgendes geschrieben: |
|
Wenn dir dabei ein Durchbruch gelingt, [...] |
| Edukir hat folgendes geschrieben: |
|
[...] will ich gerne bei dir in die Schule gehen. |
| Zitat: |
|
Der Präzisionsgriff dient der präzisen Führung der Gegenstände durch Daumen und Zeigefinger. Übrigens bei allen drei genannten Grifftechniken - man muss nur einmal genau hinschauen oder besser noch, es selbst ausprobieren. |
| Lamarck hat folgendes geschrieben: | ||
Da ist dir wohl entgangen, daß ich den vorhergehenden Beitrag nacheditiert habe, also nochmal: Es gehört zur Definition des Präzisionsgriffs, daß der gehaltene Gegenstand die Handfläche nicht berühren darf. |
| Lamarck hat folgendes geschrieben: |
|
Der Präzisionsgriff dient der präzisen Führung der Gegenstände durch Daumen und Zeigefinger. |
| Lamarck hat folgendes geschrieben: |
|
Selbstverständlich kommt der Präzisionsgriff bei der Verwendung von Wurfspeeren zum Einsatz: [...] |
| Edukir hat folgendes geschrieben: |
|
Wo kommt übrigens nach deiner Vorstellung der Präzisionsgriff her? Und der aufrechte Gang? |

| Edukir hat folgendes geschrieben: |
|
Und die "organismische Konstruktion? |
| Lamarck hat folgendes geschrieben: | ||
Einmal mehr: Es kann keine "Anpassungen an Eigenschaften" geben - s. o. Eigenschaften sind Potentiale einer organismischen Konstruktion. |
| Lamarck hat folgendes geschrieben: |
| .....
Der Anpassungsbegriff ist nicht unproblematisch. Allerdings ist er keineswegs dogmatisch, sondern axiomatisch - wie es eben so üblich ist bei Definitionen (hier: Operationale Definition). Es wird also konstatiert, dass bei 'Anpassung' die 'Keimbahn' in einer gewissen Relation mit 'Umwelt' steht. Über die Randbedingung 'Umwelt' ergibt sich somit ein Erklärungswert. Ohne diese "Ausschließlichkeit" ist der Begriff der Anpassung synonym zu einem "Ist so geworden, weil es so geworden ist". |
| Lamarck hat folgendes geschrieben: | ||
Nein. 'Verhalten' sind Freiheitsgrade organismischer Konstruktionen. Daran ändert sich auch nichts, wenn dieses Verhalten (im Extremfall die menschliche Kultur/Kulturevolution) entsprechend rückkoppeln (und wenn Du die Sache von der Keimbahn aus betrachtest, gehört Kultur ebenso zur 'Umwelt' des Menschen wie der von den Cyanobakterien produzierte Sauerstoff zur Umwelt der Cyanobakterien gehört). Apropos 'Selektionsdruck': Wer drückt denn da? |
| Lamarck hat folgendes geschrieben: | ||
D. h.: Negative Rückkopplung. |
| Lamarck hat folgendes geschrieben: | ||
Schon besser: Die Transformation von n nach n' als Anpassung der organismischen Konstruktion von der Umweltbedingung 'Wasser' an die Umweltbedingung 'Land' aus Gründen der Rekonstruktion. Schau Dir diesen einmal an: Bleibt in temporären Gewässern als letzter übrig, verfügt als Fischlarve über Außenkiemen, ist adult aufgrund asymmetrischer Lungenflügel auf atmosphärischen Sauerstoff angewiesen (Bzw.: Ist unempfindlich gegenüber Sauerstoffmangel im Gewässer) und kann als Bodenfisch auf gewisse Weise mit seinen , 'Paddeln' laufen - auch recht ausgiebig über Land. [img]Polypterus spec.[/img] ... Es gibt allerdings als Fisch noch eine ganze Reihe weiterer Strategien, um mit austrocknenden Gewässern zurechtzukommen, aber damit sollte der Anpassungsbegriff hinreichend geklärt sein. .... |
| Lamarck hat folgendes geschrieben: | ||
Hi Edukir!
Einmal mehr: Es kann keine "Anpassungen an Eigenschaften" geben - s. o. Eigenschaften sind Potentiale einer organismischen Konstruktion. Der Spielraum, wenn Du so willst. Dann kannst Du Dir überlegen, wie sich diese organismische Konstruktion über die Zeit verändern kann. Also:
Richtig: Anpassung (k) = (aufrechter Gang (Konstruktion) ∩ Präzisionsgriff (Konstruktion) ∩ n (Konstruktion) ∩ ... ) Ersteres ist finalistisches Denken mit unkritischen Vorbehalten ... . |
| Zitat: |
|
6.4 Organische Evolution Ich habe bereits angekündigt, daß man nicht darum herum kommt, sich damit zu beschäftigen, wie organische und kulturelle Evolution funktionieren, wenn man auch nur eine Chance haben will, den Verlauf der menschlichen Evolution in den letzten 1,8 MJ zu verstehen. Da der Spekulationsspielraum für diesen Zeitraum sehr hoch ist, liegt es nahe zu versuchen, ihn durch Heranziehung evolutionstheo- retischer Überlegungen einzuschränken. Die Einschränkung des Spekulations- spielraumes erkauft man dabei allerdings damit, daß die Ergebnisse der Überlegungen nun unmittelbar von der Qualität der evolutionstheoretischen Arbeitshypothesen abhängen. Sind diese Arbeitshypothesen falsch, dann gilt dies in der Regel auch für die Schlußfolgerungen über die menschliche Evolution. Aus eben diesem Grund habe ich mich bei der Betrachtung der Hominidenevolution bis zum Erscheinen des frühen Homo erectus darauf beschränkt zu untersuchen, welche Anpassungen stattgefunden haben, ohne Spekulationen darüber anzustellen, wie diese Anpassungen zustandekamen. Das Ziel der nun folgenden Überlegungen ist die Bereitstellung von Arbeits- hypothesen zu folgenden Fragestellungen: ~ Welche Eigenschaften werden im Verlauf der organischen Evolution optimiert? ~ Werden auch im Verlauf der kulturellen Evolution Eigenschaften optimiert und Wenn ja, dann welche? ~ Welchen zeitlichen Verlauf weisen die Optimierungsprozesse auf (Evolutionskinetik)? ~ In welcher Weise wirken kulturelle und organische Evolution aufeinander ein? In diesem Kapitel werden Arbeitshypothesen zur ersten und, soweit es die organische Evolution betrifft, zur dritten Frage aufgestellt. Mit der kulturellen Evolution befaßt sich das nächste Kapitel. Bei der Arbeit am SWAK-Modell der Hominidenevolution hatte ich mehrmals den Eindruck, kein wirklich neues Terrain zu betreten, sondern lediglich Darwins Spuren zu folgen, bzw. einige bereits von ihm aufgeworfene Fragen näher zu untersuchen. Auch wenn es nun darum geht, sich über einige wichtige Aspekte der Evolution klar zu werden, tut man meiner Meinung nach gut daran, Darwin im Auge zu behalten, obwohl - oder vielleicht gerade weil - er nicht die geringste Ahnung davon hatte, wie erbliche Informationen fixiert sind. Ich habe einen enormen Respekt vor Darwins Aussagen zur Evolution. Darwin hatte bei seiner Arbeit einen sehr dankbaren, wenn auch sehr aufwendigen Ansatz. Seine Theorien basieren auf einer Unmenge von Beobachtungen und haben damit einen stark deskriptiven Charakter. Da Darwins Grundannahmen ganz offensichtlich richtig waren, verfügte er darüber hinaus über ein hervorragendes Instrumentarium zur Interpretation seiner Daten. Man muß bei Darwin also 257 unterscheiden zwischen seinen Grundannahmen, zu denen die Annahme der Evolution als Wechselspiel zwischen Variation und Selektion gehört, und den Schlußfolgerungen, zu denen er durch Anwendung dieser Annahmen auf seine Beobachtungen gelangte. Seine Folgerungen waren durch eine Fülle von Fakten an die Grundannahmen gebunden und damit alles andere, als unabhängige Annahmen. Wenn Jemand aufgrund neuer Erkenntnisse (z.B. über die der Vererbung zugrundeliegenden, molekularbiologischen Prozesse) zu Aussagen gelangt, die Darwins Behauptungen widersprechen (z.B. der Behauptung, daß die der Evolution zugrundeliegenden Variationen klein sind), dann widersprechen diese Aussagen in der Regel automatisch entweder Darwins inzwischen vielfach verifizierten Grundannahmen, oder seinem empirischen Material. In beiden Fällen haben die neuen Aussagen einen schweren Stand. Dazu kommt, daß Darwin seine Evolutionstheorie selbst entwickelt hat. Als "Erfinder" dieses Gedankengebäudes hatte er einen ganz anderen Zugang zu den Daten und ein tieferes Verständnis der Zusammenhänge als irgendeiner seiner "Schüler". Wir werden meiner Ansicht nach erst dann verstehen, wie die Evolution funktioniert, wenn wir erklären können, warum sie auf der Ebene der Individuen, mit der sich Darwin auseinandergesetzt hat, gerade so funktioniert, wie Darwin es beschrieben hat. Die Beschäftigung mit den Genen, den "Elementarteilchen" der Evolution verleitet leicht dazu, ältere Erkenntnisse vorschnell in Zweifel zu ziehen, da deren Entdecker von wichtigen Aspekten der Evolution - eben von den Eigenschaften der Erbinformationsträger - keinen blassen Schimmer gehabt haben. Widersprüche der eigenen, neu entwickelten Vorstellungen zu althergebrachten Ansichten werden schnell der Unkenntnis derjenigen zugeschrieben, die diese Ansichten ihrerzeit vertraten. Moderne Vertreter der gleichen, vermeintlich überholten, Ansichten laufen Gefahr milde belächelt zu werden. Eine ähnliche Haltung legen auch manche Soziobiologen ihnen heftig widersprechenden Soziologen gegenüber an den Tag. Häufig sind jedoch gerade derartige Widersprüche Zeichen für Unzulänglichkeiten des neuen Modells und deren saubere Klärung bietet wertvolle Möglichkeiten das Modell zu verbessern. Man muß solchen Widersprüchen also unter der Prämisse nachgehen, daß etwas dran sein sollte, bis man deren Herkunft aufgeklärt hat. Statt dessen neigt man eher dazu Argumente zu suchen, die die Glaubwürdigkeit der Information, die nicht ins eigene Bild paßt, erschüttern und es einem erlauben sich vor einer ernsthaften Auseinandersetzung mit dieser Information zu drücken. Eine potentiell fruchtbare Konfrontation zwischen verschiedenen wissenschaftlichen Hypothesen entartet zu einem Stellungskampf der Wissenschaftler, der sich als Auseinandersetzung um die Rangordnung oder um Reviere eher verstehen läßt, als als gemeinsame Anstrengung im Dienste des wissenschaftlichen Fortschritts. Unser Primatenerbe bietet offensichtlich alles andere, als ideale Voraussetzungen für wissenschaftliche Betätigung. 258 Auch die klassische Thermodynamik basiert auf makroskopischen Beobachtungen. Ein tieferes Verständnis dabei eingeführter Größen, wie z.B. Wärmekapazität oder Ausdehnungskoeffizient, ergibt sich erst, wenn man die Vorgänge auf molekularem Niveau untersucht. Kein Thermodynamiker hätte deswegen jedoch eine kinetische Gastheorie akzeptiert, die in ihren Vorhersagen der klassischen Thermodynamik widerspricht. Wie funktioniert nun die organische Evolution bei höheren Säugern, zu denen ja auch der Mensch und alle seine Vorfahren im in diesem Buch betrachteten Evolutionszeitraum gehören bzw. gehörten? Die Antwort auf diese Frage ist nicht so einfach und vor allem nicht so eindeutig, wie man es vielleicht nach über hundert Jahren Evolutionsforschung erwarten würde. Während es längst keinen vertretbaren Zweifel mehr darüber geben kann, daß der Mensch ein Produkt der Evolution ist, ist die Frage, wie diese Evolution funktioniert, noch weit davon entfernt, abschließend geklärt zu sein. Ich glaube jedoch, daß man auf der Grundlage vorhandenen Wissens brauchbare Arbeitshypothesen formulieren kann mit denen sich die oben angesprochenen Fragen beantworten lassen. Es herrscht Einigkeit darüber, daß die Erbinformation in komplexen Ketten organischer Moleküle, der DNA, gespeichert wird. Diese ist aus vier verschiedenen Bausteinen aufgebaut und ihr Informationsgehalt ergibt sich aus der Reihenfolge in der diese Bausteine angeordnet sind. Abschnitte der DNA dienen als Anweisung für die Synthese von Proteinen, die wiederum verschiedenste Aufgaben übernehmen. Unter einem Gen versteht man einen Abschnitt der DNA, der eine funktionale Einheit bildet. Es gibt Strukturgene, die Proteine kodieren und Regulatorgene, die die Expression der Strukturgene steuern. Die Unterschiede im genetischen Material (Genom) zweier beliebiger Individuen einer Population sind erheblich. Das tatsächliche Ausmaß dieser Unterschiede hat die Evolutionstheoretiker seinerzeit überrascht. Im Genom eines sich geschlechtlich fortpflanzenden Lebewesens liegen Gene in zwei Versionen vor, eine stammt von der Mutter, die andere vom Vater. In etwa 10% der Fälle unterscheidet sich das eine Gen vom anderen, man spricht dann von zwei verschiedenen Allelen dieses Gens. In der Gesamtpopulation findet man zu etwa 30% der Gene unterschiedliche Allele. Zu manchen dieser Gene gibt es mehr als zwanzig verschiedene Allele im Genpool (Lewin, 1998). Diese Unterschiede in den Genotypen bilden wohl die Grundlage der beobachteten, teilweise bedeutenden Variabilität der Phänotypen bei den Individuen einer Population. Sie gehen also irgendwie mit Unterschieden im Körperbau und wohl auch im Verhalten einher. Daraus ergibt sich wiederum möglicherweise ein Vor- oder ein Nachteil für den Fortpflanzungserfolg dieser Individuen relativ zueinander und damit für das Ausmaß, in dem sich die Gene einzelner Individuen in der Population ausbreiten. Bei der sexuellen Fortpflanzung wird allerdings nicht das gesamte Genom weitergegeben, sondern jeweils nur eine Hälfte des genetischen Materials, das zeitweilig ein Genom gebildet hat, die andere Hälfte, die zur Erstellung eines neuen 259 Genoms erforderlich ist, stellt der Sexualpartner. Vom Standpunkt der Gene aus betrachtet stellt das jeweilige Genom so etwas, wie eine "Arbeitsgruppe" dar, an der sie mit dem "Ziel" teilnehmen, eigene Kopien in möglichst vielen neuen und hoffentlich ihrerseits erfolgreichen Genomen unterzubringen. Die enorme Leistungsfähigkeit dieser "Arbeitsgruppen" läßt sich an der Komplexität und den beeindruckenden Anpassungsleistungen der Individuen ablesen. Für jedes einzelne Gen eines Genoms beträgt die Chance, bei einem neuen Genom in Form einer Kopie dabei zu sein bei sexueller Fortpflanzung 50%. Im Falle asexueller Fortpflanzung, bei der das gesamte Genom an die Folgegeneration weitergegeben wird, würde sie 100% betragen. Außerdem folgt bei asexueller Fortpflanzung aus der Leistungsfähigkeit des Vorfahrengenoms mit hoher Wahrscheinlichkeit eine ebenso hohe Leistungsfähigkeit des Nachkommen- genoms. Dies trifft nur dann nicht zu, wenn das Nachkommengenom durch Mutationen nachteilig verändert wird, oder wenn die "Umwelt" sich verändert, so daß der vom Genom erzeugte Phänotyp nicht mehr so gut angepaßt ist. Die sexuelle Fortpflanzung wird daher als vom Standpunkt der Gene ausgesprochen "teure" Anpassungsleistung interpretiert, die für eine hohe Variabilität der Geno- und der Phänotypen sorgt. Diese Variabilität erhöht die Wahrscheinlichkeit, daß einige der Tochtergenome, denen die Gene ihre Kopien zur weiteren Verbreitung "anvertrauen", auch im Falle sich ändernder Umweltbedingungen zu den weiterhin erfolgreichen zählen werden. Anders ausgedrückt besagt diese Interpretation, daß sexuelle Fortpflanzung trotz der hohen für die Gene damit verbundenen Kosten so erfolgreich ist, weil die Sexualität die Möglichkeit bietet, sich schneller genetisch an veränderte Umweltbedingungen anzupassen. Das gesamte, komplexe Funktionsgefüge der sexuellen Fortpflanzung wird also als Anpassungsleistung zur Verbesserung der Evolutionsfähigkeit angesehen. Und das ergibt durchaus einen Sinn: Wenn wir uns ein zeitgenössisches, höheres Lebewesen anschauen, sagen wir mal - ohne Beschränkung der Allgemeinheit - ein Säugetier, so haben wir es bei jedem seiner Gene mit dem letzten Vertreter einer Linie von Genen zu tun, die es teilweise über Millionen von Generationen immer wieder und ohne auch nur einmal auszusetzen, geschafft haben, an Genomen "mitzuarbeiten", die der Selektion aktuelle, oft modifizierte Phänotyp-Modelle vorlegten, die besser geeignet waren den Kampf um die Fortpflanzungsressourcen unter den jeweils geltenden Bedingungen erfolgreich zu bestreiten. Zeiten schnellen Wandels, die veränderte Phänotypen bevorzugten, überstanden vor allem Genlinien, die auch in veränderten Genotypen vertreten waren. So gesehen gehören Anpassungen an die Evolutionsfähigkeit zu den potentiell vielversprechendsten Anpassungen der Evolutionsgeschichte überhaupt. Wir müssen uns also mit dem Gedanken vertraut machen, daß nicht nur Lebewesen als solche Produkte der Evolution sind. Die Evolution selbst ist ein im Laufe der Evolutionsgeschichte optimiertes Optimierungsverfahren, dessen Eigenschaften die Gene mitbestimmen und dessen sich die Gene "bedienen", um mit möglichst hoher 260 Wahrscheinlichkeit bei den erfolgreichen Genotypen der Zukunft in Form von Kopien dabei zu sein. Ich glaube, daß diese Einsicht nicht allzu weit verbreitet ist, obwohl sie in der allgemein bekannten Behauptung, die Sexualität würde dazu "dienen", die für die Evolution benötigte Variabilität bereitzustellen, enthalten ist. Der hochkomplizierte und vom Standpunkt der Fitneß sehr teure Funktionskomplex der geschlechtlichen Fortpflanzung wird also von vielen Wissenschaftlern im Prinzip als eine Anpassung an die Evolutionsfähigkeit behandelt. Dies spiegelt wider, wie hoch der evolutionäre Wert der Evolutionsfähigkeit tatsächlich ist und legt nahe nach weiteren Anpassungen dieser Art zu suchen. Es ist ziemlich widersprüchlich anzunehmen, daß höhere Organismen einerseits ein so kompliziertes, teures und mächtiges Werkzeug, wie die sexuelle Fortpflanzung, entwickelt haben, um ihre Anpassungsfähigkeit zu verbessern, daß der Verlauf der Evolution andererseits aber immer noch von zufälligen Mutationen bestimmt wird. Das entspricht in etwa der Forderung an modeme Programmierer hochintegrierte Computerprogramme durch zufällige Eingriffe in deren binären Code an neue Aufgaben anzupassen. Der Vergleich mit Computerprogrammen hinkt hierbei keineswegs. Gute Programmierer gestalten den Aufbau ihrer Programme so, daß diese mit relativ geringem Aufwand an neue, ähnliche Aufgaben angepaßt werden können, auch wenn man nicht im Voraus weiß, welche Anforderungen die neuen Aufgaben stellen werden. Die Programme werden in funktionale Einheiten (units) zerlegt, die jeweils für sich durch wohldefinierte Variablen an verschiedene Aufgaben angepaßt werden können. Die Gene sich sexuell fortpflanzender Organismen haben vielleicht ähnliche Maßnahmen ergriffen um ihre "Programme", also die Geno- und Phänotypen anpassen zu können, wie Programmierer. Hierzu ein Zitat von Gerd B. Müller (Müller, 1994): "Das Beispiel der Transformation der Extremitätenmuster zeigt, daß die Mehrzahl der phylogenetischen Struktur- und Formänderungen durch die Modulierung zellulärer und suprazellulärer Entwicklungsprozesse erreicht wird und nicht durch genetische Veränderungen auf dem Niveau von Strukturgenen. Das molekulare Repertoire der beteiligen Zellen bleibt gleich. Allerdings gibt es auch Fälle, wo Veränderungen von Strukturgenen möglicherweise einen direkten Einfluß auf die morphologische Evolution haben .... Wie die klassischen Diagramme von D' Arcy Thompson (1917) zeigen, beruhen phylogenetische Formänderungen vorwiegend auf Proportionsverschiebungen von Körperteilen. Alle dafür notwendigen relativen Vergrößerungen und Verkleinerungen von Bauelementen eines Organismus können durch die Modifikation quantitativer Entwicklungsparameter erreicht werden. Diese Parameter werden vor allem über die zeitliche Kontrolle von Entwicklungsprozessen beeinflußt. Jeder Entwicklungsprozess, ob er sich nun auf molekularer, zellulärer, oder suprazellulärer Ebene abspielt, ist durch seinen Beginn, seine Ablaufgeschwindigkeit und sein Ende definiert. Veränderungen dieser drei Variablen können im Prinzip jede erdenkliche Proportionsveränderung von Körperstrukturen bewirken. Aus diesem Grund stellt die Heterochronie - die 261 phylogenetische Veränderung von Zeitvariablen in Entwicklungsprozessen . einen der wichtigsten Mechanismen für die evolutionäre Beeinflussung der Ontogenese dar. So kann zum Beispiel ein früherer Beginn oder eine verlängerte Dauer von Zellteilungen oder eine Erhöhung der Teilungsrate die Zellenzahl und damit die Größe einer Organanlage wachsen lassen oder, im umgekehrten Fall, verringern. Dieses Konzept der Heterochronie, dessen Definition sich seit seiner Formulierung stark gewandelt hat (Gould 1977, 1988), wird immer mehr durch empirische Studien bestätigt, vor allem in der Paläontologie, aber auch in der experimentellen und vergleichenden Entwicklungsbiologie, wobei die Beispiele inzwischen eine Vielzahl von Organismen zahlreicher Taxa aus fast allen Klassen tierischer Organismen umfassen (McKinney 1988; MCKinney und McNamara 1991)." Die Evolution erfolgt also über weite Strecken durch "Modifikation quantitativer Entwicklungsparameter" die zu "relativen Vergrößerungen und Verkleinerungen von Bauelementen eines Organismus" fuhren. Ähnlich werden die Units eines Computerprogramms durch Veränderung geeignet definierter Variablen an die Lösung neuer Aufgaben angepaßt. Bei den Computerprogrammen setzt dies jedoch voraus, daß sie von vorn herein entsprechend konzipiert werden. Geniale Programme "aus einem Guß" sind zwar geeignet das konkrete Problem zu bearbeiten, für das sie entwickelt wurden, sie lassen sich jedoch kaum zur Lösung veränderter Aufgaben einsetzen. Sollte dies bei Ogranismen anders sein? Sollten diese sich in ähnlicher Weise modifizieren lassen ohne eigens dafür "entwickelt" worden zu sein? - Ich glaube nicht. Auf der anderen Seite scheint die genetische Variabilität im Genom tatsächlich auf der Akkumulation zufälliger Mutationen zu beruhen. Wie läßt sich die Funktionalität des Evolutionsprozesses auf der Ebene der Individuen mit diesen chaotischen Prozessen auf genetischer Ebene in Einklang bringen? Vermutlich ergibt sich die Funktionalität des Evolutionsprozesses nicht aus den Eigenschaften der einzelnen Gene, sondern aus der Art, wie diese Gene auf einer höheren Ebene "zusammenarbeiten". Ebenso wie sich die Funktionalität eines Phänotyps nicht auf das einzelne Gen zurückführen läßt, sondern auf dem Zusammenspiel sämtlicher Gene eines Genoms basiert, könnte die Evolutionsfähigkeit einer Population auf dem Zusammenspiel der im Genpool enthaltenen Gene basieren. Die auf der Ebene der Individuen stattfindende Selektion führt zur Funktionalität dieser Individuen, einer Funktionalität, die sich aus den Eigenschaften des Genoms ergibt und daher darin in irgendeiner Form als Information enthalten sein muß. Wir werden sehen, daß dieselbe, auf der Ebene der Individuen stattfindende Selektion im Falle der sexuellen Fortpflanzung zu einer funktionellen Variabilität der Individuen und des Genpools geführt haben könnte, die eine schnelle Anpassung der Geno- und Phänotypen ermöglicht. Der ständige Mutationsdruck wäre dann nicht die direkte Ursache der Evolution, sondern ein Aspekt der Umwelt, der bei der Entwicklung der Evolutionsfähigkeit mit berücksichtigt wurde und auf den sich das evolutionskompetente sexuelle Fortpflanzungssystem so eingestellt hat, daß sich daraus die ohnehin für die 262 Evolution benötigten genetischen Variationen im richtigen Umfang ergeben. Wie dieses evolutionskompetente sexuelle Fortpflanzungssystem (EKSF) seinerzeit zustande gekommen ist, ist eine schwierige Frage, mit der ich mich hier nicht auseinandersetzen werde. Dieser Entwicklungsprozeß liegt schon sehr weit zurück und hat womöglich die "kambrische Explosion" vor ca. 540 MJ nach sich gezogen, in der die Komplexität der jeweils komplexesten Lebewesen der Welt erheblich zunahm, wobei die "Baupläne" rezenter, höherer Lebewesen entwickelt wurden. Die von der Evolution realisierbare Komplexität eines Lebewesens hängt natürlich direkt von der Leistungsfähigkeit dieses Optimierungsverfahrens ab. Die Tatsache, daß die Lebewesen auf der Erde 3 Milliarden Jahre lang nicht über das Niveau von Einzellern hinaus kamen, könnte daher durchaus damit zusammenhängen, daß die Evolution in der Art, in der sie damals funktionierte, kein allzu leistungsfähiges Optimierungsverfahren abgab. Man sollte dabei auch nicht außer Acht lassen, daß biologische und physikalische Zeitrechnung sich nicht decken. Die biologische Zeit misst man besser in Generationsdauern - diese bezeichnen die Dauer eines Iterationsschrittes im Optimierungsverfahren Evolution. Für Mikroorganismen, deren Generationsdauer weniger als eine Stunde betragen kann, waren 3 000 MJ eine wahnsinnig lange Zeit, in der gründlich ausgelotet werden konnte, was man mit dem zur Verfügung stehenden Optimierungsverfahren erreichen konnte. Für die in diesem Buch behandelte Fragestellung ist wesentlich wichtiger, wie dieses EKSF heute funktioniert, als wie es zustande kam. Und davon kann man sich recht gut ein Bild machen: Das genetische Material eines Genpools ist sehr variabel, aber dennoch so gut aufeinander abgestimmt, daß man, wenn man aus zwei Genomen jeweils einen einfachen Satz des Erbmaterials entnimmt und zu einem neuen Genom vereinigt, dieses Genom einen Phänotyp hervorbringt, der in der Regel voll funktionstüchtig ist und ein sehr hohes Maß an Angepasstheit erkennen lässt. Genau dieser Prozess findet ständig bei der sexuellen Fortpflanzung statt und funktioniert hervorragend. Die Variationen des Phänotyps mögen in vielen Bereichen, die die Fitness kaum berühren, hoch sein, in fitneßrelevanten Bereichen sind sie gering. Das Ausmaß der Angepaßtheit hängt angesichts der enormen Variabilität des Erbguts jedoch bestimmt nicht in erster Linie vom Vorhandensein einzelner neuer, vorteilhafter Mutationen in einem Genom ab, sondern davon, wie gut die aktuelle Kombination im Genpool längst vorhandener Allele sich bewährt. Die Eigenschaften der Individuen sind im statistischen Mittel mit deren Fortpflanzungserfolg korreliert. Sie basieren auf dem Zusammenspiel der jeweiligen Allele und bleiben zum Teil auch in den Individuen der nächsten Generation erhalten. Deren Genom stammt nämlich jeweils zur Hälfte aus zwei erfolgreichen Genomen der Vorfahrengeneration und die Gene, die aus der Sicht eines der Vorfahren-Genome ausgetauscht werden, liegen immerhin in 70% der Fälle im gesamten Genpool nur in einer Version vor. Zusätzlich zu den 50% der Gene, die ohnehin zusammenbleiben, werden also mindestens 70% der 263 ausgetauschten 50% durch identische Gene ersetzt. Insgesamt entsprechen damit mindestens 85% der Kinder-Gene denen jedes der beiden Eltern. Dazu kommt noch eine unbestimmte Zahl von Genen, die in der Population zwar in mehreren Allelen vorliegen, im konkreten Fall aber durch das gleiche Allel vom anderen Elternteil ersetzt wurden und eine Vielzahl von Genen, bei denen zwar ein anderes Allel eingeführt wird, das neue Allel in seiner Wirkung auf die betrachtete Eigenschaft dem alten aber sehr nahe kommt. Eigenschaften, die auf der Kombination mehrerer konkreter Allele basieren, haben daher gute Chancen, in der nächsten Generation erhalten zu bleiben. Das stimmt mit der Beobachtung überein, daß Kinder ihren Eltern ähneln und bildet die Grundlage für die Züchtung neuer Stammlinien mit gewünschten Eigenschaften. Gehen solche Eigenschaften im Falle der natürlichen Selektion mit einem höheren Fortpflanzungserfolg einher, dann breiten sie sich gemeinsam mit der zugrunde liegenden Genkombination in der Population aus. Damit reichern sich im Laufe der Zeit in einer Population Individuen mit Eigenschaften an, die ihren Fortpflanzungserfolg maximieren. Man spricht auch davon, daß auf der Ebene der Individuen Fitneßmaximierung stattfindet. Wieso erweckt aber dieser Prozeß, soweit er sich in der Morphologie niederschlägt, den Eindruck, als würden nicht beliebige Eigenschaften verändert, sondern wohldefinierte Variablen angepaßt? Um hier eine Erklärung anzubieten, muß man etwas weiter ausholen. Wie bereits erwähnt, ist das genetische Material eines Genpools zweifellos sehr gut aufeinander abgestimmt, sonst wäre es wohl kaum möglich, von zwei Genomen jeweils die Hälfte zu entnehmen, diese Hälften zusammenzustellen und damit ein neues, voll funktionsfähiges Genom zu erhalten. Sie können ja einmal versuchen, zwei verschiedene Bücher mit der gleichen Seitenzahl einem ähnlichen Verfahren zu unterziehen, indem sie es dem Zufall überlassen, aus welchem der beiden Bücher Sie die einzelnen Seiten mit einer bestimmten Seitenzahl entnehmen. Die erste Seite könnte z.B. die erste Seite des zweiten Buches sein, der dann die zweite Seite des ersten Buches folgt und so weiter. Sie erhalten dann zwar wieder ein Buch mit der gleichen Seitenzahl und einer Unmenge von bewährten Wörtern und Sätzen - ein gutes, in sich schlüssiges Buch wird dabei nicht herauskommen. Funktionieren kann so etwas nur, wenn die beiden verwendeten Bücher in ihrem Aufbau bereits vorher auf diese Prozedur vorbereitet, - "aufeinander abgestimmt" - wurden. Auf der Ebene der Phänotypen ist gleichzeitig eine beeindruckende Funktionalität zu erkennen, die darauf hinweist, daß die Evolution als Optimierungsverfahren recht gut funktioniert. Die erfolgreichen Phänotypen kommen der optimalen Lösung der Aufgabe, den zugrunde liegenden Bauplan an die aktuellen Selektionsbedingungen anzupassen offensichtlich in der Regel sehr nahe. Diese Ansicht wird durch die Beobachtung gestützt, daß isolierte Entwicklungslinien mit ähnlichem Bauplan die gleichen Anpassungsleistungen an die gleichen Nischen 264 hervorgebracht haben. Es muß einen Grund dafür geben, daß diese getrennten Linien die gleichen Merkmalskombinationen entwickelt haben und dieser Grund besteht vermutlich darin, daß gerade diese Merkmalskombination eine optimale Anpassung an die gemeinsame Überlebensstrategie darstellt. Die Häufigkeit paralleler und konvergenter Evolution verweist daher auf die Leistungsfähigkeit der Evolution als Optimierungsverfahren und darauf, daß auf der Ebene der Phänotypen die Notwendigkeit deutlich über dem Zufall dominiert. Dazu ein Beispiel von Roger Lewin (Lewin, 1998): "Wie steht es jedoch mit den Singvögeln, einer Gruppe, der mehr als die Hälfte aller Vogelarten der Erde angehören? In Australien gibt es über 700 einheimische Arten, unter ihnen viele, die praktisch nicht von afrikanischen, europäischen oder amerikanischen Spezies unterscheidbar sind. Als man im vergangenen Jahrhundert die Klassifizierung der australischen Singvögel in Angriff nahm, hatten die europäischen Ornithologen bereits fast überall sonst auf der Welt Vögel klassifiziert. Die australischen Singvögel sahen vertrauten Arten der Alten und der Neuen Welt so ähnlich, daß es nur natürlich erschien, sie in bekannte Gruppen einzuordnen. Sibley und Ahlquist bemerken dazu, daß »viele der Konvergenzen so subtil sind, daß die wahren Verwandtschaftsbeziehungen ... sich durch anatomische Vergleiche allein wahrscheinlich nicht hätten aufklären lassen«. Aus diesem Grunde wurden zum Beispiel die Südsee-Grasmücken (Acanthiaidae) den Sylviidae (Grasmücken oder Zweigsängern) zugeordnet, die Südseeschnäpper den Muscicapidae (Sängern oder Fliegenschnäpperartigen), die Baumrutscher (Climacteridae) den Certhiidae (Baumläufern) und die Honigfresser (Meliphagedae) den nektarfressenden afro- asiatischen Nektarvögeln (Nectariniidae). Diese Klassifikation ergab morphologisch gesehen Sinn, allerdings nicht geographisch, da Australien seit mindestens 30 Millionen Jahren vom Rest der Welt getrennt ist. Die einzige Erklärung für derartige Verwandtschaftsbeziehungen wäre die wiederholte Einwanderung von Arten. anderer Kontinente gewesen - ein zwar nicht unmögliches, aber unwahrscheinliches Szenario. Überdies unterscheidet sich das von der Einwanderungshypothese implizierte Muster der Artverbreitung stark von dem der einheimischen, auf Australien beschränkten Beuteltiere, während man normalerweise, insbesondere bei räumlich begrenzter Evolution, eine geographische Übereinstimmung verschiedener Speziesgruppen erwartet. Die Daten aus der DNA-DNA-Hybridisierung stellten die bisherige Klassifikation der Vögel komplett in Frage. Sie zeigten, daß die australischen Singvögel Ureinwohner ihres Kontinents sind, brachten also die Biogeographie der Vögel mit der der Säuger in Einklang. Wie die Beuteltiere haben sich auch die Singvögel Australiens an ökologische Nischen angepaßt, die den Nischen in anderen Regionen der Erde gleichen, und ähneln ihren dort heimischen Gegenstücken infolge konvergenter Evolution, wobei der Grad der Ähnlichkeit oft erstaunlich ist." Da die meisten Individuen einer Population sehr gut angepaßt sind, sind starke Veränderungen für die Fitness relevanter Eigenschaften eines Kindes im Vergleich zu den Eltern fast immer mit einer deutlichen Reduktion der Fitness verbunden. 265 Allele, die in verschiedenen Genomen zu deutlich unterschiedlicher Ausprägung solcher Eigenschaften fuhren würden, würden regelmäßig nach erfolgter Rekombination Ausschuß produzieren. Die Genome, an denen sie mitarbeiten, würden eben aufgrund dieser Mitarbeit im statistischen Mittel weniger erfolgreich sein, was letztendlich zur Eliminierung solcher Allele aus dem Genpool führen würde. Daher gibt es in einem Genpool nur selten solche Allele, wenn sie infolge von Mutationen auftreten, dann werden sie in der Regel sehr bald wieder heraus selektiert. Die 70% der Gene, die in einem Genpool nur in einer einzigen Variante vorliegen, dürften auf Mutationen recht empfindlich mit deutlich verändertem Einfluss auf fitneßrelevante Eigenschaften des Phänotyps reagieren. Aus diesem Grund haben sich auch keine Variationen dieser Gene im Genpool festsetzen können. Die Behauptung, daß die Darwin’sche Evolution, die die Bedeutung der Selektion für den evolutionären Fortschritt betont, die Annahme nahelegt, daß gerade Mutationen, die starken Einfluß auf den Phänotyp ausüben, sich besonders schnell im Genpool ausbreiten sollten, ist falsch. Für ein Optimierungsverfahren, das so aufgebaut ist, wie die Evolution, ist mit einer starken Abhängigkeit seiner Leistungsfähigkeit vom Ausmaß der Variation in den einzelnen Iterationsschritten zu rechnen. Entsprechendes kennt man bei mathematischen Optimierungsverfahren und es ist bekannt, daß eine zu starke Variation dazu führt, daß das Optimierungsverfahren nicht mehr konvergiert, d.h. das Optimum wird nicht mehr gefunden. Warum dies so ist, kann man sich anschaulich recht einfach klarmachen: Nehmen wir einmal an, wir wollten uns mit einem der Evolution strukturell ähnlichen Verfahren einem bestimmten, uns unbekannten Punkt A möglichst stark nähern. Wir hätten eine Meßvorrichtung, die uns zu jedem untersuchten Punkt X den Abstand von A liefert. Nun würden wir eine Anzahl beliebiger X untersuchen und denjenigen mit dem geringsten Abstand zu A zum Arbeitspunkt des ersten Iterationsschrittes machen. Um diesen Arbeitspunkt würde nun eine Schar von neuen Punkten X erzeugt, die in verschiedenen Richtungen in einem Abstand von 1-3 Metern von ihm liegen. Jeder dieser X würde nun auf seinen Abstand von A untersucht und der nächstgelegene zum neuen Arbeitspunkt für den nächsten Iterationsschritt bestimmt. Mit diesem Verfahren würden wir uns zweifellos zuverlässig bis auf eine Entfernung in der Größenordnung von einem Meter an A annähern können. Sollten wir dann jedoch zufällig einen Arbeitspunkt erhalten, der sich in nur 40 cm Abstand von A befindet, dann würde die Erzeugung einer Schar neuer Punkte mit einem Abstand von 1-3 Metern vom Arbeitspunkt im nächsten Iterationsschritt zwangsläufig dazu fuhren, daß kein einziger dieser Punkte näher an A liegt, als 60 cm. Für eine Variation um 3 m ergibt sich dabei sogar ein Mindestabstand von 2,6 Metern, eine solche Variation hätte daher von vorn herein keine Chance, den nächsten Arbeitspunkt zu stellen. Eine neue punktschar um den Arbeitspunkt in 60 cm Entfernung könnte A wiederum nicht näher, als bis auf 40 cm kommen, u.s.w .. Das Verfahren würde um den gesuchten Punkt springen, ohne sich ihm weiter 266 nähern zu können. Daraus erkennt man, daß eine zuverlässige Annäherung an A im Bereich von Zentimetern voraussetzt, daß auch die Variation in der gleichen Größenordnung liegt. Die ideale Variationsgröße für einen Iterationsschritt hängt stark davon ab, wie weit man noch vom Optimum entfernt ist. Ist man diesem nahe, so führen nur noch kleine Variationen weiter. Die natürliche Auslese, der bei der Evolution eine entscheidende Bedeutung zukommt, entspricht der Auswahl eines Arbeitspunktes aus einer Schar neu erzeugter Punkte in diesem gedachten Optimierungsverfahren. Im Falle der Evolution sich sexuell fortpflanzender Lebewesen haben wir es, wie bereits erwähnt, mit sehr gut angepaßten Lebewesen zu tun - also mit Arbeitspunkten, die bereits sehr nahe am gesuchten Punkt liegen. Daraus abgeleitete, neue Punkte, haben nur dann eine Chance zu Arbeitspunkten des nächsten Iterationsschrittes zu werden, wenn sie sich aus einer geringen Variation der Arbeitspunkte ergeben - die Kinder werden nur dann eine hohe Fitneß aufweisen, wenn ihre Eigenschaften nicht zu stark von denen ihrer Eltern abweichen. Die natürliche Selektion sorgt daher dafür, daß genetische Variationen mit starkem Einfluß auf die Eigenschaften der Individuen nicht Fuß fassen können. Nur neue Allele, die geringen (funktional kaum schädlichen) Einfluß auf wichtige (fitneßrelevante) Eigenschaften ausüben, setzen sich im Genpool fest. Dies entspricht zwar den entsprechenden Aussagen der Neutralitätstheorie der Evolution, steht jedoch keinesfalls in einem Widerspruch zur Darwinistischen Evolutionstheorie. Das gestaltgebende Prinzip der Evolution, der entscheidende Vorgang, der funktionale Komplexität erst möglich macht, ist weiterhin die Selektion. Die Neutralitätstheorie betrifft lediglich die Entstehung genetischer Variabilität und beleuchtet damit nur einen Teilaspekt der Evolution. Einen Teilaspekt zudem, der bei der Entwicklung neuer Eigenschaften, der wir uns nun zuwenden, vermutlich nur eine untergeordnete Rolle spielt. Von der Annahme, daß sich im Genpool vor allem Variationen festsetzen, die nur geringen Einfluß auf fitneßrelevante, funktionelle Aspekte auf der Ebene der Individuen besitzen, bis zu der Beobachtung, daß Evolution vor allem durch Variation quantitativer Parameter erfolgt, ist nur noch ein kleiner Schritt. Die prinzipielle Funktionsfähigkeit eines Organs hängt in der Regel nur schwach oder gar nicht von dessen Größe ab. Größenänderungen fuhren zwar zu Veränderungen der Leistungsfähigkeit, der "Wirtschaftlichkeit" und anderer optimierbarer Aspekte, sie haben aber nur in Ausnahmefällen Einfluß auf die Funktionsfähigkeit. Damit sind quantitative Parameter, wie z.B. die Größen einzelner Bauelemente die idealen Werkzeuge um einen gegebenen Bauplan an die Erfordernisse verschiedener Aufgaben anzupassen, ohne dessen Funktion deutlich zu beeinträchtigen - es gibt genügend Beispiele aus der Technik, in denen gerade so verfahren wird. Genetische Variationen, deren Auswirkung sich auf quantitative Aspekte der Individualentwicklung beschränken sind damit aber - eben weil sie die Funktionsfähigkeit der Phänotypen nur selten in Frage stellen und daher oft vom Standpunkt der Gesamtfitneß zu den "kleinen" Variationen gehören - auch 267 diejenigen, die sich am ehesten der Neutralitätstheorie zufolge im Genpool ausbreiten können sollten (wenn man einmal von den tatsächlich neutralen Variationen absieht, die überhaupt keinen Einfluß auf den Phänotyp ausüben). Es reichern sich also im Falle der sexuellen Fortpflanzung genetische Variationen im Genpool an, die zu einer hohen Variabilität gerade der Eigenschaften führen, mit deren Anpassung man zweckmäßigerweise auf der Ebene der phänotypischen Evolution an veränderte Anforderungen reagieren sollte. Diese Variationen stellen wiederum das Handwerkszeug dar, das vom EKSF beim Optimierungsprozeß herangezogen wird. Daß dieses maßgeschneiderte "Handwerkszeug" im Ernstfall bereits zur Verfügung steht, ist vermutlich das Erfolgsgeheimnis der sexuellen Fortpflanzung. Einerseits ermöglicht es die schnelle Anpassung an eine sich verändernde Umwelt - diesen Vorteil der sexuellen Fortpflanzung haben wir bereits angesprochen. Andererseits erklärt es aber auch, warum Lebewesen, die sich sexuell fortpflanzen, höhere Komplexität erreichen konnten. Bei komplexen Organismen müssen sehr viele Eigenschaften sehr fein aufeinander abgestimmt werden. Dazu bedurfte es im Verlauf derer Evolution einer Unmenge geeigneter, kleiner Variationen von der Art, wie sie bei sich sexuell fortpflanzenden Organismen in großer Zahl vorliegen und immer wieder neu bereitgestellt werden. Die Möglichkeit komplexere Organismen zu entwickeln und damit in bis dahin noch nicht besetzte Nischen vorzudringen stellt einen weiteren Vorteil der sexuellen Fortpflanzung dar und hat wohl - vielleicht sogar entscheidend - zum Erfolg dieses Fortpflanzungssystems beigetragen. Bei komplexen Lebewesen mit stark ausdifferenzierten und spezialisierten Körperteilen üben Veränderungen des Bauplans in der Regel sehr starken Einfluß auf die Funktionalität und damit auch die Fitneß aus. Mutationen, die solche Veränderungen nach sich ziehen, haben daher in aller Regel keine Chance, sich im Genpool festzusetzen. Daher stellt die Evolution komplexer Organismen seit geraumer Zeit lediglich eine Folge von Interpretationen des gleichen Grundmusters dar. Der Pudel ist so gesehen eine extreme Interpretation des Wolfes, der Mensch im Großen und Ganzen eine extreme Interpretation des Schimpansen (genaugenom- men sind beide zwei verschiedene Interpretationen des gleichen Grundmusters). Es ist bisher nicht gelungen im Gehirn des Menschen auch nur eine funktionale Einheit aufzuspüren, über die der Schimpanse nicht - zumindest im Ansatz - ebenfalls verfügt. Die Evolution verläuft über weite Strecken durch Neuabstimmung vorhandener funktionaler Einheiten, wodurch im Extremfall auch ganz neue Eigenschaften erzeugt werden können, wie es z. B. die Sprachfähigkeit des Menschen ist. Auch hier werden Einheiten verwendet, die anscheinend allesamt auch schon beim Schimpansen vorhanden sind, allerdings im Verlauf der menschlichen Evolution enorm verändert werden mußten. Die Spezies, die man jeweils beobachten kann, sind lediglich Interpretationen innerhalb der Möglichkeiten, die ihnen durch ihre jeweilige Kombination funktionaler Einheiten und verfügbarer Variablen vorgegeben werden. Welche Interpretation jeweils vorherrscht, hängt von den Umweltanforderungen 268 und von der vorhergegangenen Entwicklung ab. Die vorhergegangene Entwicklung spielt eine große Rolle, weil das Optimierungsverfahren mit dem Namen Evolution in der Regel nur dann funktioniert, wenn die Variationen klein sind. Mit echten Entwicklungssprüngen ist daher im Verlauf der genetischen Evolution kaum zu rechnen. Punktmutationen, die zu erheblichen Veränderungen des Phänotyps führen sind zwar jederzeit möglich - konstruktiv sind sie jedoch mit an Sicherheit grenzender Wahrscheinlichkeit nicht. Es kommt zwar vor, daß zwei deutlich unterschiedliche Ausprägungen des Phänotyps sich vom Standpunkt der Fitneß aus betrachtet nicht unterscheiden und nebeneinander in der Population halten können - dies sind jedoch eher Ausnahmefälle. Ausnahmefälle allerdings, die in der wissenschaftlichen Diskussion eine sehr große Rolle gespielt haben, eine wesentlich größere jedenfalls, als im tatsächlichen Evolutionsverlauf. Die Beobachtung zeigt, daß die Evolution der Lebewesen, soweit man darunter einen phänotypischen Wandel versteht, in Schüben erfolgt. Lange Zeiträume phänotypischer Stabilität wechseln sich mit kurzen Phasen dynamischer Veränderungen ab. Die genetische Variation wird dagegen kontinuierlich erzeugt, in vielen Bereichen des Genoms zeigt sie geradezu die Eigenschaften einer molekularen Uhr, so daß man zum Beispiel aus der Anzahl der sich in verschiedenen Entwicklungslinien angesammelten Mutationen auf den Verzweigungszeitpunkt dieser Linien zu schließen vermag. Wie wir oben gesehen haben, führt die genetische Evolution aber vermutlich gerade bei den Eigenschaften zu einer hohen Variabilität innerhalb der Population, aus denen sich die Evolution auf der Ebene der Phänotypen bevorzugt zusammensetzt. Auf die meisten dieser Eigenschaften übt eine ganze Reihe von Genen einen Einfluß aus und die genetische Evolution führt dazu, daß gerade diese Gene im Genpool einer Population in Form - mitunter sehr vieler - verschiedener Allele auftreten. Eine bestimmte Größe - z.B. die Länge eines Oberschenkelknochens - ergibt sich aus dem summarischen Einfluß aller entsprechenden Allele eines Genoms. Man kann sich den Genpool als Lebensraum dieser Allele vorstellen, in dem z.B. alle Allele, die für einen längeren Oberschenkelknochen "plädieren" miteinander konkurrieren. Ihre "Bevölkerungsdichte" hängt dabei davon ab, wie lang der Oberschenkelknochen unter den gegebenen Umweltbedingungen tatsächlich sein sollte. Diese für die Fitneß des Individuums entscheidende Größe wird durch die Selektion diktiert - wie sie im Einzelnen realisiert wird, liegt im "Ermessensspielraum" der Gene. Werden von der Selektion lange Knochen verlangt, dann wachsen die Populationen der "Langknochen-Allele im Genpool, währen die Populationen der "Kurzknochen"-Allele kleiner werden. Nur gelegentlich kommt es wohl dazu, daß einzelne Allele vollständig aus dem Genpool eliminiert werden. Meistens bleibt die Variabilität des Genpools wohl erhalten - es ändert sich lediglich die Häufigkeiten des Auftretens einzelner Allele. Dies führt dazu, daß phänotypische Evolution über weite Strecken im Prinzip umkehrbar verläuft. Derartige Umkehrungen der Evolution, wie z.B. Reaktivierungen alter Muster, sind mehrfach beobachtet worden, ohne daß man bisher verstanden hat, 269 wie es dazu kommen kann. Ein Beispiel, mit dem wir in diesem Buch bereits zu tun hatten, sind die Überaugenwülste des Homo erectus. Sie sind ein ursprüngliches Merkmal der Menschenaffen, das im Verlauf der Hominidenevolution reaktiviert wurde. Es wurde höchste Zeit sich an die Hominidenevolution zu erinnern, denn ich bin in diesem Kapitel bereits deutlich von der Fragestellung abgewichen, mit der sich dieses Buch befassen soll. In diesem Kapitel sollten lediglich zwei Arbeitshypothesen aufgestellt werden, die die organische Evolution betrafen. Einmal ging es darum, welche Eigenschaften im Verlauf der organischen Evolution optimiert werden, zum anderen ging es um den zeitlichen Verlauf solcher Optimierungsprozesse. In beiden Fällen interessiert uns lediglich die Ebene der phänotypischen Evolution. Bei der ersten Arbeitshypothese stehe ich im Einklang mit den meisten Wissenschaftlern und hätte sie auch einfach in Form eines Zitats bereitstellen können: Die organische Evolution führt zur Maximierung der Fitneß der Individuen, die man neuerdings auch gerne als Darwin-Fitneß bezeichnet. Das Einzige, was man für die Formulierung dieser Hypothese braucht, ist die Erkenntnis, daß genetische Informationen über die Fortpflanzung weitergegeben werden und variabel sind. Für eine brauchbare Formulierung der zweiten Hypothese ist es dagegen hilfreich, wenn man eine brauchbare Vorstellung davon hat, wie genetische Information strukturiert ist und zu den Veränderungen des Phänotyps in Beziehung steht. Entscheidend für meine Arbeitshypothese ist dabei, daß Populationen aufgrund ihrer genetischen Variabilität im Prinzip "evolutionskompetent" sind. Es ist nicht unbedingt erforderlich, davon auszugehen, daß tatsächlich eine Evolution der Evolutionsfähigkeit stattgefunden hat (insofern bin ich mit meinen Ausführungen tatsächlich über das Ziel hinausgeschossen). Wenn die bei der phänotypischen Evolution verwendeten Variationen in erster Linie durch sexuelle Rekombination bereits im Genpool vorhandenen, variablen genetischen Materials zustande kommen (und das ist gegenwärtig Lehrmeinung), dann können Populationen praktisch "aus dem Stand" mit Anpassungen auf veränderte Selektionsbedingungen reagieren. Sie müssen nicht erst auf geeignete Mutationen warten. Sie sind damit im Prinzip "evolutionskompetent". Damit liegt die Annahme nahe, daß Populationen auf veränderte Selektionsbedingungen im Prinzip ähnlich reagieren, wie Regelsysteme auf Veränderungen der Eingangsgrößen. Eine Reaktion mit besonderer evolutionstheoretischer Relevanz ist dabei die sogenannte "Sprungantwort" . Die Sprungantwort ist der Verlauf, den der geregelte Wert (Ausgangswert) über die Zeit aufweist, wenn man den Sollwert am Eingang des Regelsystems sprunghaft ändert. Wenn sich Populationen ähnlich verhalten, wie ein Regelsystem, dann würden sie in ähnlicher Weise auf sprunghafte Änderungen der Selektions- 270 bedingungen reagieren. Solche können sich durch schnelle Veränderungen der Umwelt oder durch Verhaltensänderungen ergeben. Im SWAK-Modell spielen vor allem Verhaltensänderungen eine große Rolle, diese stellen aber oft ihrerseits eine Reaktion auf eine sich ändernde Umwelt dar. Bild 18: Der Verlauf der Sprungantwort eines einfachen Regelsystems, wie er qualitativ im Verlauf der Evolution vermutlich häufig vorkommt. Die horizontale Achse ist die Zeitachse, auf der vertikalen Achse läßt sich der Verlauf der Regelgröße ablesen. Der Sollwert, den das Regelsystem einstellen soll, wurde zum Zeitpunkt t=0 von 0 auf I verändert. Daß eine "Sprungantwort" mit dem in Bild 18 dargestellten, qualitativen Verlauf für evolutionäre Szenarien tatsächlich einen Sinn ergibt, sollen folgende Überlegungen illustrieren. Betrachten wir z.B. ein "Entwick- lungsvorhaben", wie den Übergang zum aufrechten Gang am Beginn der Hominidenevolution. Eingeleitet wurde dieser Prozeß - soweit das SWAK-Modell - durch eine Verhaltensänderung, die ihrerseits als Antwort auf klimatische Veränderungen gesehen wird. Die Vorfahrenpopulation bestand aus Knöchel- gängern, die fähig waren aufrecht zu gehen, wenn auch nicht besonders gut. lnfolge der Verhaltensänderungen wurden die Selektionsbedingungen - gemessen an evolutionären Zeiträumen - sprunghaft verschoben. Leistungsfähigkeit im Knöchelgang wurde nun geringer, Leistungsfähigkeit beim aufrechten Gang höher bewertet. Die Population reagierte darauf sofort mit einer Verschiebung der Eigenschaften, die zur Verbesserung des aufrechten Ganges auf Kosten des Knöchelganges führte. Solange der aufrechte Gang noch nicht allzu gut funktionierte, boten sich natürlich viele Ansatzpunkte ihn deutlich zu verbessern. Geringfügige Verschiebungen in der Geometrie z.B. des Kniegelenkes konnten zu deutlicher Verbesserung dieser Eigenschaft führen. Am Beginn der Entwicklung wurden daher schnell Fortschritte erzielt. Mit Annäherung an das neue Optimum wurde es jedoch immer schwieriger den inzwischen bereits deutlich verbesserten aufrechten Gang noch weiter zu verbessern. Die Entwicklungsgeschwindigkeit nahm im Verlauf des Anpassungs- prozesses kontinuierlich ab, da es einerseits schwieriger wurde, weitere Verbesserungen zu erzielen und andererseits kleinere Verbesserungen auch mit geringerem Selektionsvorteil einhergingen und sich daher auch langsamer in der Population ausbreiteten. Am Ende folgten "Feinabstimmungen", die nur noch wenig zur Leistungsfähigkeit beim aufrechten Gang beitrugen, dafür aber - gemessen am 271 dynamischen Beginn des Anpassungsvorganges - noch geraume Zeit in Anspruch nahmen. Die für die Dynamik des Anpassungsvorganges relevanten Aspekte dieses Beispiels entwickeln auch bei anderen "Entwicklungsvorhaben" in gleicher Weise ihre Wirkung. Es ist also damit zu rechnen, daß die "Sprungantwort" eines EKSF strukturell ähnlich verläuft, wie wir es aus der Regelungstechnik für manche Regelsysteme kennen. Schaut man sich unter diesem Gesichtspunkt den Verlauf der Evolution an, soweit er uns aus Fossilien bekannt ist, dann gewinnt man den Eindruck, daß die Evolution sich vor allem aus solchen Sprungantworten zusammensetzt, die sporadisch - in langen Zeiträumen phänotypischer Stabilität eingebettet - auftreten. Hier finden wir auch die von Gerhard Roth geforderte Erklärung dafür, daß das Gehirnvolumen bei unseren Vorfahren drei Phasen schnellen Wachstums aufweist. Es handelt sich um drei "Sprungantworten", die auf entsprechende Änderungen der Selektionsbedingungen hinweisen. Das Gehirn zeigt dabei lediglich die gleiche Evolutionskinetik, die wir auch beim Körperbau beobachten können - z.B. beim Übergang zum Homo erectus. Wenn wir nun an die Abbildung von Schrenk zurückdenken (Bild 16, Kap. 6.1), dann wird auch klar, was ich gegen den Verlauf, den er für die Entwicklung der Gehirnstruktur und der kognitiven Leistungen postuliert, einzuwenden habe. Seine Kurven zeigen die falsche Krümmung, was der Annahme entspricht, daß die Weiterentwicklung dieser Eigenschaften sich mit der Zeit immer weiter beschleunigt habe. Wie ich jedoch oben ausgeführt habe, ist für einen Entwicklungsschub innerhalb der organischen Evolution im Gegenteil mit einem schnellen Beginn und einem langsamen Auslaufen zu rechnen und infolgedessen mit dem umgekehrten Krümmungsverlauf der Entwicklungskurve. Oft leiten solche Entwicklungsschübe im Verlauf der Evolution radiative Speziationen ein. Auch dies ist verständlich, denn eine Verschiebung des Anpassungsmusters kann die Möglichkeit eröffnen, neue Nischen zu erobern, vor allem, wenn sie mit der Entwicklung neuer Fähigkeiten einhergeht. Eine solche Sicht des Evolutionsverlaufs war meiner Meinung nach auch schon zu Darwins Zeiten, unter Zugrundelegung seiner Evolutionstheorie, naheliegend. Darwin stützte sich bei seiner Theoriebildung ja auf die beobachtete phänotypische Variabilität in Populationen, er wußte, daß diese groß ist und jederzeit für Anpassungsprozesse herangezogen werden konnte. Immerhin unterhielt er intensive Beziehungen zu Züchtern und führte selbst Züchtungsexperimente durch. In solchen Experimenten wird die "evolutionäre Kompetenz" von Populationen offensichtlich, da sich durch willkürliche Selektion neue Eigenschaften "produzieren" lassen. Ihm war auch bekannt, wie gut einzelne Lebewesen an ihre Lebensweise angepaßt sind, schließlich war er ja in einem Umfeld aufgewachsen, in dem begeisterte Naturforscher immer neue Arten beschrieben und in derer Funktionalität Beweise 272 für das Wirken eines allmächtigen Schöpfers sahen. All dies hätte eigentlich eine Sicht nahe legen sollen, der zufolge die meisten Lebewesen bereits optimiert sind, die phänotypische Evolution im Wesentlichen stagniert und es nur noch in Einzelfällen infolge veränderter Selektionsbedingungen zu Anpassungsprozessen kommt. Man hätte also auf der Grundlage der Darwinschen Evolutionstheorie sehr wohl brauchbare Aussagen über die Evolutionskinetik machen können. Stattdessen ging man in der Anfangszeit der Evolutionstheorie von einem kontinuierlichen Fortschritt in der Evolution aus. Vermutlich lagen dieser Interpretation vor allem der Zeitgeist und das altehrwürdige, anthropozentrische Gedankengebäude des scala naturae zugrunde. Der Zeitgeist war von einem starken Fortschrittsglauben durchtränkt, denn die Gesellschaft befand sich mitten in einer Phase dynamischer, technologischer Entwicklung. Dieser Fortschrittsglaube wurde auf die Evolutionstheorie projiziert, obwohl für diese die technologische Entwicklung des neunzehnten Jahrhunderts nicht im Geringsten von Belang war. In der scala naturae wurde andererseits eine natürliche Rangordnung unter allen Lebewesen postuliert, an deren Spitze natürlich der Mensch stand. Solcher Ballast, der wohl letztendlich auf unser Primatenerbe zurückzuführen ist, prägt auch heute noch unser Denken und lag solchen Wortschöpfungen, wie z.B. den "höheren Säugetieren" zugrunde. Wollte man das dem Menschen schmeichelnde Konzept der scala naturae beibehalten, so lag es nahe, anzunehmen, daß die niederen Tiere in ihrer Evolution einfach noch nicht so weit entwickelt waren wie die Höheren. Wenn sie jedoch noch nicht voll entwickelt waren, dann besaßen sie noch Spielraum zur weiteren Verbesserung, die vermutlich ständig stattfand. Hierbei spielte dann auch das nichtdarwinistische Konzept eines den Lebewesen innewohnenden "inneren Entwicklungsdranges" in der Anfangszeit der Evolutions- theorie eine Rolle. Die wenigsten Wissenschaftler stimmten damals Darwins mechanistischem Konzept einer Evolution in Form eines Wechselspiels zwischen zufälliger Variation und natürlicher Selektion zu. Der Bruch, der hier mit den Traditionen und vor allem mit den religiös geprägten Weltanschauungen vollzogen werden mußte, war zu groß, als daß eine saubere Auswertung der Implikationen der Evolutionstheorie möglich gewesen wäre. Die frühen Vorstellungen zur Evolutionskinetik, die später mit der Darwinschen Evolutionstheorie assoziiert wurden und im Widerspruch zum Fossilbefund stehen, wurden also gar nicht aus der Evolutionstheorie abgeleitet. Vielmehr wurde die Evolutionstheorie mit herrschenden Vorurteilen in einem neuen Gedankengebäude oberflächlich in Einklang gebracht. In wissenschaftlich ähnlich unsauberer Weise wurde die Evolutionstheorie von rassistischen und sozialdarwinistischen Strömungen mißbraucht. Diese stellen aber wiederum ihrerseits Aspekte der kulturellen Evolution dar, mit deren Gesetzmäßigkeiten wir uns nun beschäftigen werden. 273 |
output generated using printer-friendly topic mod. Alle Zeiten sind GMT + 1 Stunde